জীবদেহে অন্তঃকোষীয় বার্তাবাহক হিসেবে ক্যালসিয়ামই সম্ভবত অগ্রজ
- প্রকাশ: ০২:৫৮:৫২ অপরাহ্ন, শনিবার, ২ জুলাই ২০২২
- / ৮৮৪ বার পড়া হয়েছে
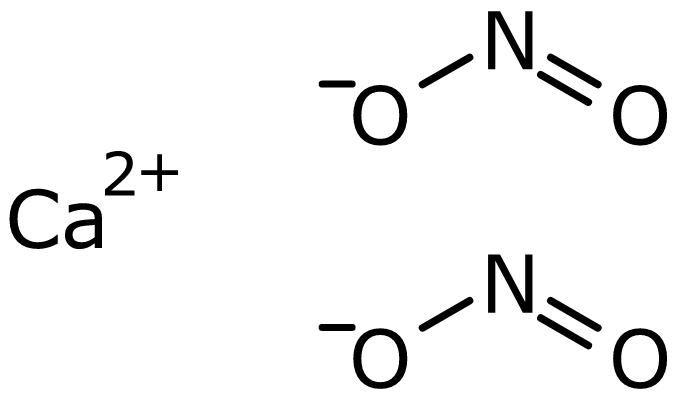
ক্যালসিয়াম
পৃথিবী হচ্ছে সূর্যের দেহজাত। ধরা হয় ৪.৬ বিলিয়ন বছর আগে পৃথিবীর জন্ম এবং তার কিছু সময় (প্রায় ৫০ মিলিয়ন বছর) পরেই পৃথিবীর গা থেকে বেরিয়ে এসেছে চাঁদ নামক উপগ্রহটি। সূর্যের মধ্যে যে-সব উপাদান রয়েছে তার মধ্যে হাইড্রোজেন (৯১.২ শতাংশ) ও হিলিয়াম (৮.৭ শতাংশ) হচ্ছে প্রধান, আর বাকি যেসব উপাদান (অক্সিজেন, কার্বন, নাইট্রোজেন, সিলিকন, ম্যাগনেসিয়াম, নিয়ন, আয়রন, সালফার, ক্যালসিয়াম ইত্যাদি) সেখানে পাওয়া যায় তা পরিমানে বেশ নগণ্য, ০.১ শতাংশেরও কম। যেহেতু সূর্য-পৃষ্ঠের ১০,০০০ ডিগ্রি সে. প্রখর তাপে পরমাণুগুলো অত্যন্ত গরম, তাই তারা ছুটে বেড়ায় খুব দ্রুত বেগে। প্রায়শই তারা সংঘর্ষে লিপ্ত, ফলে পরমাণুর বাইরের কক্ষপথ থেকে দু-একটা ইলেকট্রন সহজেই ছিটকে পড়া অসম্ভব কিছু নয়। একটি নিরপেক্ষ ক্যালসিয়াম পরমাণুর প্রোটন ও ইলেট্রনের সংখ্যা হলো ২০টি। দুটি ইলেক্ট্রন হারিয়ে সে অবস্থান নিলো ডাইভালেন্ট ধনাত্মক আয়নে। একটি নবজাত পৃথিবী ছিল বর্তমান পৃথিবীর থেকে সম্পূর্ণ আলাদা। পৃথিবী সৃষ্টির প্রথম যুগে গলিতবস্তু ঠান্ডা হতে শুরু করলো তখন জলীয়বাষ্প আর আঙ্গারিক গ্যাস (কার্বন ডাইঅক্সাইড-CO2)র সৃষ্টি হলো। পৃথিবীর তাপ কমে গিয়ে জোলো বাষ্প সমুদ্রে বিস্তৃতি লাভ করলো। সে সময়ে বায়ুমণ্ডলের প্রধান উপাদান ছিল আঙ্গারিক গ্যাস (CO2), প্রায় ৯৫ শতাংশ, এবং মুক্ত অক্সিজেনের অস্তিত্ব বেশ নগণ্য, ছিল না বল্লেই চলে। পরবর্তী পর্যায়ে বাতাসে কিছু অক্সিজেন ও নাইট্রোজেন দেখা দিয়েছিল। বায়ুমণ্ডলীয় আঙ্গারিক গ্যাস, অক্সিজেন ও নাইট্রোজেনের প্রাকৃতিক উৎসের মধ্যে রয়েছে আগ্নেয়গিরির গ্যাসীয়-উদগিরণ। এছাড়াও, অক্সিজেনের উদ্ভব হয়েছিল যখন প্রারম্ভিক প্রাক-ক্যামব্রিয়ান (pre-cambrian) যুগে আঙ্গারিক গ্যাসের অঙ্গার (carbon) পদার্থ নিয়ে গঠিত সাইয়ানোব্যাকটেরিয়া (Cyanobacteria) অর্জন করলো সালোকসংশ্লেষণ বৈশিষ্ট্য (প্রায় ৩.৫ বিলিয়ন বছর আগে) এবং মুক্তি দিলো অক্সিজেনকে।
আমরা যদি বর্তমান মঙ্গল গ্রহের সঙ্গে তুলনা করি, তাহলে সেখানেও দেখবো গ্রহটি ঢাকা রয়েছে ৯৫% আঙ্গারিক গ্যাসে। অন্যান্য গ্রহেও হয়ত CO2-র আধিক্য রয়েছে। সময়ের সাথে সাথে আদি পৃথিবীর বায়ুমণ্ডলের কার্বন ডাই অক্সাইড পরিবর্তিত হয়েছিল ক্যালসিয়াম কার্বনেট (CaCO3) বা চুনাপাথরে। বাইকার্বোনেট উৎপাদিত হয় চুনাপাথর, জল এবং কার্বন ডাই অক্সাইডের মধ্যে বিক্রিয়ার মাধ্যমে, মুক্ত হয় ক্যালসিয়াম আয়ন। উদাহরণস্বরূপ: CaCO3+CO2+H2O ⇌ Ca2++2HCO−3। ফলস্বরূপ, বায়ুমণ্ডলে CO2-এর উপস্থিতি কমে আসলো।
পৃথিবীর যে প্রাপনীয় ক্যালসিয়াম তা, অন্যান্য পরমাণুর মতোই, পৃথিবী সাথে করে নিয়ে এসেছে তার সৃষ্টির সময়ে। শুধু পৃথিবীতে হয়েছে তার গাঠনিক ও ভূতাত্ত্বিক পরিবর্তন। ক্যালসিয়াম প্রকৃতিতে দেখা যায় যৌগিক অবস্থায়, যেমন কার্বনেট, সালফেট, ফ্লোরাইড, সিলিকেট এবং বোরেট আকারে।
একটি বিষয় লক্ষণীয়, ক্যালসিয়াম যেখানেই অবস্থান করছে সেখানেই তার স্থান পঞ্চম। পৃথিবীপৃষ্ঠে অক্সিজেন, সিলিকন, অ্যালুমিনিয়াম এবং লোহার পরেই তার অবস্থান; সমুদ্রের পানিতে সোডিয়াম, ক্লোরাইড, ম্যাগনেসিয়াম এবং সালফেটের পরেই হচ্ছে এই দ্রবীভূত আয়ন; এবং মানবদেহে মৌলের দিক হতে ক্যালসিয়ামের অবস্থানও পঞ্চম (অক্সিজেন, কার্বন, হাইড্রোজেন এবং নাইট্রোজেনের পরের স্থান হচ্ছে ক্যালসিয়াম), তবে ধাতুর দিক থেকে প্রথম। ক্যালসিয়ামের গুরুত্ব দেখা দিল সকল জীব ও মানবদেহের একটি অপরিহার্য উপাদান হিসেবে। আদি ও প্রকৃতকোষের অভ্যন্তরে যে ক’টা সংকেতবাহী (messenger molecule) অণু বা আয়নের উদ্ভব হয়েছে তার মধ্যে ডাইভালেন্ট ক্যালসিয়াম আয়ন হচ্ছে অগ্রজ।
আমাদের দেহের প্রায় ৯৬ শতাংশ (ভর অনুপাতে) চারটি মূল উপাদান দিয়ে তৈরি: অক্সিজেন (৬৫%), কার্বন (১৮.৫%), হাইড্রোজেন (৯.৫%) এবং নাইট্রোজেন (৩.৩ শতাংশ)। এই উপাদানগুলো দেহের গুরুত্বপূর্ণ অণুগুলোতে পাওয়া যায়, যার মধ্যে রয়েছে পানি, প্রোটিন, ডিএনএ ও আরএনএ (RNA), এবং অন্যান্য জৈব অণুতে। মৌলের দিক হতে পঞ্চম অবস্থানে থাকা ক্যালসিয়ামের পরিমাণ হচ্ছে ১.৫%, যার অধিকাংশ (৯৯%)-ই যৌগিক অবস্থায় গঠন করে দেহের হাড় এবং দাঁত। আর বাকিটুকু (১ শতাংশ) রয়ে গেল মুক্ত বার্তাবাহী আয়ন হিসেবে, যার শারীরিক ক্রিয়াকলাপ পরিচালনায় অসীম গুরুত্ব নিয়েই লেখা আমার এই নিবন্ধ। দেহে পেশি সংকোচন, হরমোন নিঃসরণ, রক্ত জমাট বাঁধা, স্নায়ু-সিনাপ্সে নিউরোট্রান্সমিটার মুক্তি ও সংকেত প্রবাহ এবং হৃদস্পন্দন নিয়ন্ত্রণে ক্যালসিয়াম একটি অপরিহার্য উপাদান। মস্তিষ্ক থেকে দেহের সমস্ত অঙ্গে কোষীয় সংকেত বহনে দায়িত্ব পেলো যে কয়টি আয়ন বা অণু তাদের মধ্যে ক্যালসিয়াম অন্যতম। সমস্ত কোষগুলোকে একে অপরের সাথে সংকেত সংবহনের মাধ্যমে যোগাযোগ স্থাপন করে দিলো ক্যালসিয়াম।
আদি যুগে বায়ুমণ্ডলে যেখানে আঙ্গারিক গ্যাসের ছিল প্রাধান্য তা কালক্রমে হ্রাস পেতে পেতে বর্তমানে তার ঘনত্ব এসে দাঁড়িয়েছে মাত্র ৪১৫ পিপিএম (PPM: parts per million) বা ০.০৪ শতাংশে। অপরপক্ষে সে স্থান দখল করেছে নাইট্রোজেন (৭৮%) ও অক্সিজেন (২১%)। এই গ্যাসীয় বিপ্লবের পেছনে যে প্রক্রিয়ার অবদান সব থেকে বেশি তা হলো সালোকসংশ্লেষণ, যার সূত্রপাত হয়েছিল সায়ানোব্যাকটেরিয়া বা নীল-সবুজ শৈবাল (যদিও এটি শৈবাল নয়) সৃষ্টির মাধ্যমে। বর্তমানে, বায়ুমন্ডলীয় আঙ্গারিক গ্যাসের ঘনত্ব সমগ্র উদ্ভিদজগতের সালোকসংশ্লেষন দ্বারা নিয়ন্ত্রিত।
আমরা খাদ্য থেকে ক্যালসিয়াম গ্রহণ করি খনিজ আকারে। দাঁত-অস্থির গঠন ছাড়াও কোষের বিভিন্ন বিপাকীয় ক্রিয়াকলাপের জন্য অন্ত্রে ক্যালসিয়ামের শোষণ অপরিহার্য। খাদ্য থেকে গৃহিত ক্যালসিয়ামের মাত্র ৩৫-৪০ শতাংশ অন্ত্রে শোষিত হয়। বস্তুতঃ খাদ্যে উপস্থিত ফাইটিক অ্যাসিড (phytic acid) ক্যালসিয়াম এবং অন্যান্য খনিজকে অদ্রবণীয় করে তোলে ও শোষণে বাধা দেয়। অন্ত্রের ঝিল্লি জুড়ে ক্যালসিয়াম শোষণ হচ্ছে মূলতঃ ভিটামিন-ডি (1,25(OH)2D3)-নির্ভর। এছাড়া, ভিটামিন ডি-অসম্পৃক্ত পথেও ক্যালসিয়ামের কিছুটা শোষণ হয়। অধিকন্তু, প্যারাথাইরয়েড ও প্রোল্যাক্টিন হরমোনের প্রভাবেও ক্যালসিয়াম শোষণের মাত্রাকে বৃদ্ধি করে। ভিটামিন-ডি এর উদ্দীপনায় TRP (Transient Receptor Potential)-রিসেপ্টর (এক ধরণের ক্যালসিয়াম চ্যানেল) ও ক্যালসিয়াম-বাইন্ডিং প্রোটিন ‘ক্যালবিনডিন’ (calbindins)-এর মাধ্যমে ক্যালসিয়াম শোষিত হয় রক্তপ্রবাহে। TRP-R ও ক্যালবিনডিন অন্ত্রের এপিথেলীয় কোষ (এন্টেরোসাইট)এ অবস্থান করে। অন্ত্রের ডুডেনাম (duodenum) ক্যালসিয়াম শোষণের প্রধান অঞ্চল, যদিও ক্ষুদ্রান্তের অন্যান্য অঞ্চল এবং কোলন থেকেও ক্যালসিয়াম শোষিত হয়। কিডনিতেও দেখা গেছে প্রায় ৬০% থেকে ৭০% ক্যালসিয়াম প্রক্সিমাল টিউবুলে নিষ্ক্রিয়ভাবে পুন:শোষিত হয়।
সংকেত-সংবহনে ক্যালসিয়াম পরিবহনের সুবিধার্ধে কোষাভ্যন্তরে রয়েছে কিছু ক্যালসিয়াম-বহনকারী বা ক্যালসিয়াম-বাইন্ডিং প্রোটিন (CaBP)। এই প্রোটিনগুলো ক্যালসিয়াম-নিয়ন্ত্রিত কোষীয় সংকেত সংবহনে অংশগ্রহণ করে। সমস্ত প্রকৃতকোষীয় (ইউক্যারিওটিক) জীবে Ca2+-নির্ভর সংকেতের প্রাথমিক মধ্যস্থতাকারী প্রোটিন হলো ক্যালমোডুলিন (calmodulin, CaM), যা একটি অন্তঃকোষীয় Ca2+ রিসেপ্টর হিসেবে কাজ করে। বলা বাহুল্য, ব্যাক্টেরিয়ায় ক্যালমোডুলিনের উপস্থিতি নেই। তবে, ক্যালমোডুলিন-অনুরূপ কিছু প্রোটিন রয়েছে, যার মাধ্যমে আদিকোষীয় ব্যাকটেরিয়াও Ca2+-নির্ভর সংকেত ব্যবহার করে ও তাদের কোষাভ্যন্তরে Ca2+-এর পরিমাণ (৮০-১০০ nM) কঠোরভাবে নিয়ন্ত্রণে রাখে। ব্যাকটেরিয়ায় বিভিন্ন প্রক্রিয়ার সাথে জড়িত রয়েছে ক্যালসিয়াম, যেমন কোষের গঠন, গতিশীলতা, ডিএনএ সংশ্লেষণ, কোষ বিভাজন, জিনের অভিব্যক্তি নিয়ন্ত্রণ ইত্যাদি। এমনকি সায়ানোব্যাকটেরিয়া (যা সম্ভবত সবচেয়ে প্রাচীন জীব যা আজও বেঁচে আছে)তেও Ca2+ এর প্রয়োজন হয় যৎসামান্য।
ক্যালমোডুলিন প্রোটিনের নির্দিষ্ট ডোমেন বা অঞ্চল রয়েছে যেখানে আবদ্ধ হয় ক্যালসিয়াম। কালমোডুলিনের সক্রিয়তা আসে শুধুমাত্র ক্যালসিয়ামের সাথে আবদ্ধতায়। CaBP-এর প্রধান কাজ হচ্ছে কোষের সাইটোপ্লাজমে মুক্ত ক্যালসিয়াম (Ca2+) এর পরিমাণ নিয়ন্ত্রণ করা, পরিবহনে সহযোগিতা করা ও ক্যালসিয়ামের ভারসাম্যতা (হোমিওস্ট্যাসিস) বজায় রাখা। কালমোডুলিন একটি আদর্শ ক্যালসিয়াম-বাইন্ডিং প্রোটিন। ক্যালসিয়ামের সাথে আবদ্ধ হয়ে কালমোডুলিন বিভিন্ন লক্ষ্যবস্তুতে, যেমন কাইনেজ (kinase), বিশেষকরে CaMK (Ca2+/calmodulin-dependent protein kinase), ও ফসফেটেজ (phosohatase) এনজাইমকে উদ্দীপিত করে। CaMK-IV ট্রান্সক্রিপশন ফ্যাক্টর CREB (cAMP response element-binding protein)-কে ফসফোরিলেশনের মাধ্যমে প্রত্যক্ষভাবে কিছু গুরুত্বপূর্ণ জিনের, যেমন BDNF (Brain-derived neurotrophic factor), BCl-2 (B-cell lymphoma 2), GAD67(Glutamate decarboxylase), parvalbumin (ক্যালসিয়াম-বাইন্ডিং প্রোটিন), TXNIP (Thioredoxin-interacting protein) প্রকাশকে বৃদ্ধি করে। অনুরূপভাবে, ক্যালসিয়ামের উদ্দীপনায় ক্যালসিনুরিন (calcineurin), একটি ক্যালসিয়াম- এবং ক্যালমোডুলিন-নির্ভর সেরিন/থ্রিওনাইন প্রোটিন ফসফেটেজ, ইমিউন সিস্টেমের টি-কোষ (T-cells)গুলোকে সক্রিয় করে। পেশি সংকোচন, স্নায়ু সংকেত পরিবহন, দৃষ্টিশক্তি নিয়ন্ত্রণসহ অনেক জিনের প্রকাশকেও কালমোডুলিন নিয়ন্ত্রণ করে। একটি গবেষণায় দেখা গেছে, Ca2+/কালমোডুলিন-উদ্দীপ্ত অ্যাডিনাইলেট সাইক্লেজ (ADCY1) এনজাইম সির্কাডিয়ান (circadian) ছন্দে cAMP (Cyclic Adenosine Monophosphate) তৈরি করে, রাত্রিতে বেশি ও দিনে কম। অর্থাৎ Adcy1 প্রতিলিপন (transcription) ও Ca2+/কালমোডুলিন-উদ্দীপ্ত ক্রিয়াকলাপ জৈবিক ঘড়ি (biological clock) দ্বারা নিয়ন্ত্রিত (Chaurasia S, Haque R, Pozdeyev N, et al., J Neurochem. 2006, 99(4):1142-1150)। কালমোডুলিন ছাড়াও আরও CaBP রয়েছে, যেমন CaBP4, ক্যালবিন্ডিন ও ক্যালসেকোয়েস্ট্রিন (Calsequestrin)।
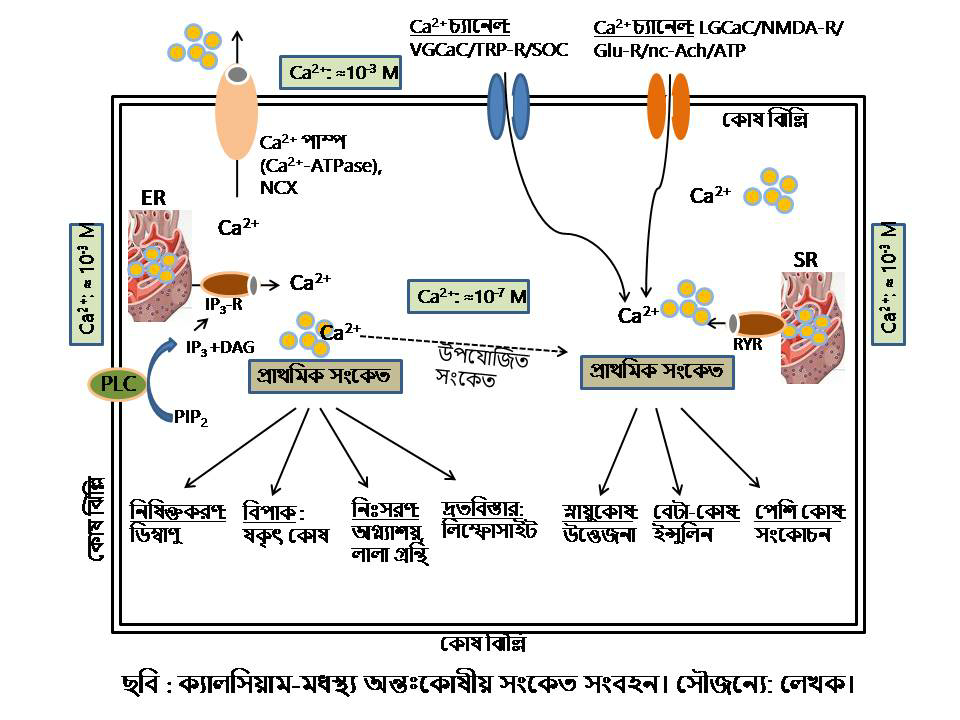
প্রকৃতকোষে সংকেতবাহী আয়ন হিসেবে ক্যালসিয়ামের আগমন একটি মাইলফলক। বিশ্রামকালীন বা স্থিতিশীল অবস্থায় ক্যালসিয়ামের কোষাভ্যন্তরস্থ ঘনত্ব থাকে ≈10−7M, যখন তার বহিঃস্থ ঘনত্ব থাকে ≈10−3M)। ফলে, কোষের এপার-ওপার একটি Ca2+ ঘনত্বের ইলেক্ট্রোকেমিক্যাল গ্রেডিয়েন্ট তৈরি হয়, যা কোষাভ্যন্তরে ক্যালসিয়াম আনয়নে একটি প্রেরণার উৎস। ইউক্যারিয়োট কোষঝিল্লি জুড়ে ক্যালসিয়াম পারাপারের যে প্রবেশপথগুলোর অবস্থান তার কতকগুলো (Ca2+ পাম্প, যা কাঠামোগতভাবে ইউক্যারিয়োটিক P-টাইপ Ca2+ পাম্পের মত, এবং Ca2+/H+ ও Na+/Ca2+ এক্সচেঞ্জার) এমনকি ব্যাক্টেরিয়াতেও দেখা যায়। এছাড়াও, ব্যাকটেরিয়ায় রয়েছে কিছু আদি আয়ন চ্যানেল ও অ-প্রোটিনধর্মী Ca2+ আয়ন চ্যানেল। ছত্রাক থেকে স্তন্যপায়ী প্রাণী পর্যন্ত বিস্তৃত ইউক্যারিওটিক জীবে অন্তঃকোষীয় সংকেত সংবহনের একটি গুরুত্বপূর্ণ অংশ হচ্ছে Ca2+ আয়ন-মধ্যস্থ সংকেত, যা কোষের পৃষ্ঠ থেকে কোষের অভ্যন্তরে সংকেত প্রেরণ করে ও Ca2+-নিয়ন্ত্রিত জিনগুলির অভিব্যক্তি বৃদ্ধি বা প্রতিহত করে। কোষের অন্যান্য বার্তাবাহক হচ্ছে সাইক্লিক নিউক্লিওটাইড (cAMP ও cGMP), ইনোসিটল ট্রিসফসফেট (IP3) এবং ডায়াসিলগ্লিসেরল (DAG)। কোষঝিল্লি ঘেঁষা এনজাইম ফসফোলাইপেজ C (PLC)-এর অনুঘটনে লিপিড PIP2 (Phosphatidylinositol 4,5-bisphosphate) ভেঙে তৈরি হয় IP3 ও DAG।
কোষপৃষ্ঠে অবস্থানরত সেকেলে একটি সার্বজনীন প্রোটিন হলো Ca2+-ATPase বা ক্যালসিয়াম পাম্প (calcium pump), যা ইলেক্ট্রোকেমিক্যাল গ্রেডিয়েন্টের উপর ভিত্তি করে সক্রিয়ভাবে কোষের অতিরিক্ত Ca2+ আয়ন কোষের বহিঃস্থ স্পেসে নির্গত করে। পেশি কোষের সারকোপ্লাজমিক রেটিকুলামের ঝিল্লিতেও রয়েছে এই ধরণের ক্যালসিয়াম পাম্প। অন্তঃকোষীয় ক্যালসিয়ামের ঘনত্ব সর্বদা স্থিতিশীল অবস্থায় (≈ ১০০ nM) রাখতে কোষের এটি একটি আণবিক কৌশল। যেহেতু ATPase গ্রুপের প্রোটিন, তাই তারা এই কাজটি করার জন্য শক্তি আহরণ করে ATP অণু ভেঙে। প্রতিটি ATP অণুর আদ্রবিশ্লেষে দুটি ক্যালসিয়াম আয়ন পরিবহন করা হয়। ক্যালসিয়াম পাম্প ছাড়াও ঝিল্লিবন্ধ সোডিয়াম-ক্যালসিয়াম এক্সচেঞ্জার (Na+/Ca2+exchanger, NCX) প্রোটিনটিও অন্তঃকোষীয় Ca2+ ঘনত্বের প্রধান নিয়ামক হিসেবে কাজ করে। সোডিয়াম-ক্যালসিয়াম এক্সচেঞ্জার কোষ থেকে ক্যালসিয়াম অপসারণের এই কাজটি করে এক অভিনব কায়দায়। NCX-র একটি নীতি হলো যে ৩:১ অনুপাতে তিনটি Na+ আয়ন কোষে প্রবেশ করে এবং একটি Ca2+ আয়ন কোষ থেকে বেরিয়ে যায়। এই দেয়া-নেয়ায় কোষাভ্যন্তরে ক্যালসিয়ামের ঘনত্ব ও ভোল্টেজের মাত্রা নিয়ন্ত্রণে এক্সচেঞ্জারটি যথেষ্ট অবদান রাখে।
কালানুক্রমে সুকেন্দ্রিক জীবে কোষঝিল্লি জুড়ে ক্যালসিয়ামের চলাচল জটিল হয়েছে উত্তোরোত্তর। সিস্টেমের জটিলতার বড় একটি কারণ হলো কোষগুলোর নির্দিষ্ট Ca2+ সংকেত প্রক্রিয়ার সাথে অন্তঃকোষীয় অঙ্গাণু (বিশেষকরে এন্ডোপ্লাজমিক রেটিকুলাম, ER)গুলোর বিকাশ ঘটেছে। ভিন্ন এক ধারায় রেটিকুলামের ঝিল্লি জুড়ে Ca2+ প্রবেশ/প্রস্থানের জন্য তাদের নিজস্ব কিছু ব্যবস্থা রয়েছে। উদাহরণস্বরূপঃ উল্লেখ করা যেতে পারে IP3 রিসেপ্টরের (IP3-R) কথা, যা উদ্দীপ্ত হয় IP3 (inositol 1,4,5-trisphosphate) বার্তাবাহী অণুর দ্বারা। ফলশ্রুতিতে, রেটিকুলামে সঞ্চিত ক্যালসিয়াম মুক্ত হয় সাইটোপ্লাজমে। মাইটোকন্ড্রিয়া নামক অঙ্গাণুতেও রয়েছে সে ব্যবস্থা, সেখানেও সঞ্চিত থাকে অতিরিক্ত কালসিয়াম। এই অঙ্গাণুদ্বয়কে ক্যালসিয়াম-ভান্ডার বলা হলে তা অত্যুক্তি হবে না।
ক্যালসিয়াম পাম্প ও সোডিয়াম-ক্যালসিয়াম এক্সচেঞ্জার ছাড়াও কোষের ঝিল্লিতে উদ্ভব হয়েছে কঠোরভাবে নিয়ন্ত্রিত কিছু আয়ন চ্যানেল। আয়ন হলো নিট বৈদ্যুতিক আধানযুক্ত কণিকা, পরমাণু বা অণু। কোষঝিল্লিতে আবদ্ধ পরিবাহীসক্ষম এই আয়ন চ্যানেলগুলো হলো প্রোটিনধর্মী, যা কোষের উত্তেজনা নিয়ন্ত্রণে প্রধান ভূমিকা পালন করে। এই চ্যানেলগুলো কার্যত: প্রায় সব ধরণের কোষেই পাওয়া যায়, যদিও স্নায়ুতন্ত্র ও পেশিতন্ত্রে এই চ্যানেলগুলো পরিব্যাপ্ত এবং এদের গুরুত্ব অসীম। এই পরিবাহী চ্যানেলগুলো মূলতঃ তিন গ্রুপে বিভক্ত: ভোল্টেজ-গেটেড, বহিঃকোষীয় লাইগ্যান্ড-গেটেড এবং অন্তঃকোষীয় লাইগ্যান্ড-গেটেড আয়ন চ্যানেল। এই চ্যানেলগুলো অবাধ নয়, কঠোরভাবে আয়ন-নির্দিষ্ট গেট দ্বারা নিয়ন্ত্রিত। অর্থাৎ, নির্দিষ্ট আয়ন-উদ্দীপকের সাড়া পেলেই শুধুমাত্র চ্যানেলগুলো খোলে, আবার বন্ধ হয়, ঠিক যেন স্লুস (sluice) গেটের মত। তাই, এই চ্যানেলগুলোকে পারিভাষিক শব্দ হিসেবে যথার্থই ‘আয়নকপাট’ও বলা যেতে পারে, যার মধ্য দিয়ে নির্দিষ্ট অণুগুলো কোষের ঝিল্লি জুড়ে পরিবাহিত হয়।
ভোল্টেজ-গেটেড ক্যালসিয়াম চ্যানেল (voltage-gated calcium channel, VGCaC)গুলো ঝিল্লি জুড়ে ভোল্টেজের তারতম্যে নিয়ন্ত্রণ করে ক্যালসিয়াম পরিবহন। VGCaC একটি আয়ন চ্যানেল, যা শুধুমাত্র ক্যালসিয়াম আয়ন পারাপারকে নিয়ন্ত্রণ করে। তবে, মাঝে-মধ্যে তাদের অল্প পরিমাণ সোডিয়াম আয়নকেও ছাড় দিতে দেখা গেছে, কিন্তু VGCaC-র মধ্য দিয়ে সোডিয়াম আয়নের থেকে ক্যালসিয়াম আয়নের প্রবেশাধিকার হাজার গুণ বেশি। এই প্রোটিন চ্যানেলগুলো উত্তেজনা-সঞ্চারী (excitatory) কোষ (যেমন পেশি, গ্লিয়া কোষ, নিউরন)গুলোর ঝিল্লিতে আবদ্ধ। এইসব উত্তেজনা-সৃষ্টিকারী কোষগুলোর মেমব্রেন পোটেনশিয়াল (কোষঝিল্লি বরাবর বৈদ্যুতিক বিভব) যখন স্থিতিশীল বা রেস্টিং অবস্থায় (resting membrane potential, RMP) থাকে তখন VGCaC বন্ধ থাকে। প্রসঙ্গত উল্লেখ্যঃ একটি স্নায়ুকোষাভ্যন্তরে যখন বাইরের তুলনায় আধান থাকে ঋণাত্মক, অর্থাৎ নেতিবাচক –৭০ মিলিভোল্ট (mV) বা তার আশপাশ, তখন সেটিকে বলা হয় RMP। একটি VGCaC সক্রিয় হয় যখন কোষের মেমব্রেন পোটেনশিয়াল RMP-র থেকে কম ঋণাত্মক বা ধনাত্মক আধানের হয় (যেমন -৫০ থেকে +৪০ mV)। কোষের এই আয়নভিত্তিক মেরুকরণকে বলা হয় ডিপোলারাইজেশন (depolarization), যখন ক্যালসিয়াম চ্যানেলগুলো খুলে যায় এবং Ca2+ আয়ন সাইটোপ্লাজমে প্রবেশ করে। কোষের এই বৈদ্যুতিক বিভবকে বলা হয় অ্যাকশন পোটেনশিয়াল। ফলস্বরূপঃ ক্যালসিয়াম চ্যানেলের সক্রিয়তায় পেশি সংকোচন, নিউরনের উত্তেজনা, হরমোন ও নিউরোট্রান্সমিটার নিঃসরণ ইত্যাদি সংঘটিত হয়। স্তন্যপায়ী প্রাণীদের মধ্যে VGCaC পরিবারের দশটি সদস্য রয়েছে যাদের সংশ্লেষণে সংকেত প্রদানের জন্য DNA-তে দশটি পৃথক জিনের উদ্ভব হয়েছে, তবে এগুলো প্রধানত পাঁচটি ভাগে বিভক্ত: L-type, N-type, P/Q-type, R-type, এবং T-type, তম্মধ্যে L-type ক্যালসিয়াম চ্যানেল অত্যন্ত গুরুত্বপূর্ণ। বলা বাহুল্য, ব্যাক্টেরিয়ার Ca2+ চ্যানেলের ভোল্টেজ-নির্ভরতা প্রকৃতকোষীয় লো-ভোল্টেজ-অ্যাক্টিভেটেড T-type ক্যালসিয়াম চ্যানেলের মত।
কোষে ক্যালসিয়ামের আধিক্যে দেহে উচ্চ রক্তচাপ বা হাইপারটেনশন হওয়ার সম্ভাবনা অনেক বেশি। এল-টাইপ (L-type) ক্যালসিয়াম চ্যানেল প্রতিবন্ধক বা অ্যান্টাগোনিস্ট (যেমন, amlodipine [dihydropyridine]), যার কার্যকারিতা দীর্ঘস্থায়ী উচ্চ রক্তচাপ বা হাইপারটেনশন ও ‘এনজাইনা পেকটোরিস’ চিকিৎসায় ব্যবহৃত হয়। রক্তনালিকার মসৃণ পেশি টিস্যুতে ডিপোলারাইজেশন (depolarization) আবহের সৃষ্টি হওয়ায় কোষে ক্যালসিয়াম ও সোডিয়াম আয়নের প্রবেশের মাধ্যমে দেখা দেয় হাইপারটেনশন। এল-টাইপ ক্যালসিয়াম চ্যানেল-প্রতিবন্ধক অ্যামলোডিপিন রক্তনালিকার মসৃণ পেশি ও হৃদপেশিতে ডিপোলারাইজেশন-জনিত নির্দিষ্ট অ্যাকশন পটেনশিয়াল (action potential)কে বাধা দেয়। ফলস্বরুপ রক্তনালীর (বিশেষকরে করোনারি ধমনী) প্রসারণের মাধ্যমে পর্যায়ক্রমে রক্তচাপকে স্বাভাবিক পর্যায়ে নিয়ে আসে।
অপরপক্ষে, লাইগ্যান্ড-গেটেড ক্যালসিয়াম চ্যানেল (LGCaC)গুলো আয়োনোট্রপিক রিসেপ্টর। এই চ্যানেলগুলোও প্রোটিনধর্মী। জীবজগতে লাইগ্যান্ড-গেটেড চ্যানেলগুলোর উপস্থিতি বেশ পুরোনো। এক ধরণের পেন্টামেরিক রিসেপ্টর সায়ানোব্যাকটেরিয়ায় প্রোটন-তাড়িত চ্যানেল হিসেবে বিদ্যমান, যা মেরুদন্ডী প্রাণীদের ক্ষেত্রে অ্যাসিটিলকোলাইন, γ-অ্যামিনোবুটারিক অ্যাসিড (GABA), গ্লাইসিন বা সেরোটোনিন জাতীয় লাইগান্ড দ্বারা নিয়ন্ত্রিত চ্যানেলের সমরূপ। প্রকৃতকোষীয় জীবে অন্তঃকোষীয় অঙ্গাণু, বিশেষকরে এন্ডোপ্লাজমিক রেটিকুলাম (ER) এবং মাইটোকন্ড্রিয়ায় বেশ কিছু বৈচিত্র্যময় বৈশিষ্ট্য যুক্ত হয়েছে LGCaC ও VGCaC পরিবারে, জটিল থেকে জটিলতর হয়েছে তাদের কার্যবিধি। প্রোটিস্টা (যেমন অ্যামিবা, প্যারামেসিয়াম ইত্যাদি) গ্রুপ থেকে শুরু করে স্তন্যপায়ী সমস্ত প্রাণীদের মধ্যে এন্ডোপ্লাজমিক রেটিকুলাম ও মাইটোকন্ড্রিয়ার ঝিল্লিতে আবদ্ধ LGCaC-রিসেপ্টর (যেমন IP3 রিসেপ্টর)র উপস্থিতিতে কোষাভ্যন্তরে ক্যালসিয়ামের নিয়ন্ত্রণ এক ভিন্ন মাত্রায় গিয়ে দাঁড়ায়। নিঃসন্দেহে, সবকিছুর সমন্বয়েই কোষে বার্তাবাহী আয়ন হিসেবে নির্ভুলভাবে সংকেত সংবহন করা ক্যালসিয়ামের জন্য একটি চ্যালেঞ্জ হয়ে দাঁড়ায়।
বার্তাবাহী অণু, হরমোন বা নিউরোট্রান্সমিটারের মতো লাইগ্যান্ডের আবদ্ধতায় (LGCaC) চ্যানেলগুলো উম্মুক্ত হয় ও Ca2+ আয়ন-কে ঝিল্লির মধ্য দিয়ে যাবার অনুমতি দেয়। লাইগ্যান্ড-গেটেড ক্যালসিয়াম চ্যানেলের মধ্যে রয়েছে IP3 রিসেপ্টর ও রাইয়ানোডিন (Ryanodine, RyR ) রিসেপ্টর। এছাড়াও, অনেক নিউরোট্রান্সমিটার, যেমন গ্লুটামেট (Glu), GABA, NMDA (N-Methyl-D-aspartic acid), নিকোটিনিক অ্যাসিটিলকোলাইন (nc-Ach) এবং ATP-নিয়ন্ত্রিত LGCaC-গুলো সার্বজনীন ধনাত্মক আয়নভেদযোগ্য- তবে, চ্যানেলগুলোর মধ্য দিয়ে বিশেষ করে Ca2+ আয়নের প্রবেশ অন্যান্য আয়নগুলোর থেকে অনেক বেশি। এই রিসেপ্টরগুলোর আধিক্য কেন্দ্রীয় স্নায়ুতন্ত্রে। অন্তঃকোষীয় এন্ডোপ্লাজমিক রেটিকুলাম (ER) বা (পেশি কোষের) সার্কোপ্লাজমিক রেটিকুলাম (SR)-এ সঞ্চিত থাকে কোষের অতিরিক্ত ক্যালসিয়াম তা পূর্বেই বলা হয়েছে। প্রয়োজনের তাগিদে সঞ্চিত ক্যালসিয়াম দেহের অস্থি বা ER/SR থেকে ক্যালসিয়াম নিঃসৃত হয় কোষাভ্যন্তরে একটি নিয়মের মাধ্যমে। ER/SR ঝিল্লির সাথে যুক্ত দুই ধরনের চ্যানেল (RyR এবং IP3 রিসেপ্টর)র সক্রিয়তা আসে যথাক্রমে স্বয়ং Ca2+ আয়ন ও IP3 বার্তাবাহী অণু দ্বারা। পেশি সংকোচন, মেমব্রেন পোটেনশিয়াল, স্নায়ুতন্ত্রে ও পেশিকোষে সংকেত সংবহন ইত্যাদি নিয়ন্ত্রিত হয় LGCaC-এর মাধ্যমে ক্যালসিয়ামের মধ্যস্থতায়। আরও এক ধরণের ক্যালসিয়াম-প্রবেশযোগ্য ঝিল্লিবন্ধ LGCaC চ্যানেল, যা P2X রিসেপ্টর নামে অভিহিত, উদ্দীপ্ত হয় ATP লাইগ্যান্ডের মাধ্যমে। P2X শ্রেণীর পূর্বসূরি আয়নোট্রপিক পিউরিনোসেপ্টর পাওয়া যায় প্রোটোজোয়া জীবদের মধ্যে। এই চ্যানেলগুলি দেখা যায় স্তন্যপায়ী প্রাণীদের স্নায়ুতন্ত্রে, মাইক্রোগ্লিয়ায়, ম্যাক্রোফেজে, জরায়ুর এন্ডোমেট্রিয়ামে এবং রেটিনায়। এছাড়া, স্তন্যপায়ীদের প্রজননতন্ত্রে আবির্ভুত হয়েছে বিশেষ এক ধরণের Ca2+-তাড়িত LGCaC (PKD2 পরিবার), যা স্ত্রী-প্রজনন ট্র্যাক্টে ডিম্বাণুকে খুঁজে নিতে শুক্রাণুর একান্ত সহায়ক।
ক্যালসিয়াম দেহের অস্থি ও দাঁত গঠন ছাড়াও অন্তঃকোষীয় সংকেত সংবহন পথগুলোতে একটি গুরুত্বপূর্ণ ভূমিকা পালন করে, যেখানে Ca2+ আয়ন দ্বিতীয় স্তরের বার্তাবাহক (the second messenger molecule/ion) হিসেবে কাজ করে, নিউরন থেকে নিউরোট্রান্সমিটার মুক্তিতে, ইমিউন সিস্টেমের বি-কোষ ও টি-কোষের উদ্দীপনায়, পেশি কোষের সংকোচনে এবং নিষিক্তকরণে। (জেনে রাখা ভালো, প্রথম স্তরের বার্তাবাহী অণু হলো লাইগান্ড বা উদ্দীপক এবং তৃতীয় স্তরের বার্তাবাহক হচ্ছে ট্রান্সক্রিপশন ফ্যাক্টর, যা ডিএনএ-তে অবস্থিত জিনগুলিতে সরাসরি আবদ্ধ হয়ে তাদের প্রকাশকে বৃদ্ধি বা প্রতিহত করে)। অনেক এনজাইমের কোফ্যাক্টর হিসেবেও ক্যালসিয়াম আয়ন প্রয়োজন, যার মধ্যে বেশ কয়েকটি রক্ত ক্লটিং ফ্যাক্টরও রয়েছে। দেহের প্রত্যেকটি কোষে, এমনকি এককোষীয় জীবেও, ক্যালসিয়ামের ভারসাম্য বজায় রাখতে ভিন্নমুখী দুটি কৌশলের উদ্ভব হয়েছে। সংকেত এবং কোষের প্রকার ভেদে এক পথে যেমন অন্তঃকোষীয় মুক্ত Ca2+ এর প্রবাহ বৃদ্ধি পায় লাইগান্ড- ও ভোল্টেজ-গেটেড ক্যালসিয়াম চ্যানেলের মাধ্যমে, তেমনি অন্য পথে ক্যালসিয়াম বহির্গামী প্রক্রিয়ায় ক্যালসিয়াম ATPase পাম্প এবং অ্যান্টি-পোর্ট সিস্টেম (NCX)-এর মাধ্যমে অতিরিক্ত মুক্ত ক্যালসিয়াম কোষের বাইরে অপসারিত হয় অথবা এন্ডোপ্লাজমিক রেটিকুলাম অঙ্গাণুতে তা সঞ্চিত রাখে। লক্ষ্য একটাই, কোষাভ্যন্তরে Ca2+ কে ন্যানোমোলার (≈ ১০০ nM) পরিসরে রেখে তার স্থিতিশীলতা বজায় রাখা ও অ্যাপোপটোসিসের মত অপমৃত্যুর হাত থেকে নিজেকে রক্ষা করা। ক্যালসিয়াম আয়ন (Ca2+) জীবকোষের শারীরবৃত্ত ও জৈব রসায়নে এক অভাবনীয় অবদান রেখে চলেছে যুগ থেকে যুগান্তর। কত যে লাইগান্ড-উদ্দীপ্ত খবর সে অবিরত বয়ে নিয়ে যায় কোষঝিল্লি থেকে ডিএনএ পর্যন্ত, তার খবর আমরা ক’জনেই বা রাখি! এই নিবন্ধটি লিখতে গিয়ে কবি সুকান্ত ভট্টাচার্য-র একটি জনপ্রিয় কবিতা মনে পড়লো:
“রানার ছুটেছে তাই ঝুম্ঝুম্ ঘণ্টা বাজছে রাতে
রানার চলেছে, খবরের বোঝা হাতে,
রানার চলেছে রানার!
রাত্রির পথে পথে চলে কোনো নিষেধ জানে না মানার।
দিগন্ত থেকে দিগন্তে ছোটে রানার-
কাজ নিয়েছে সে নতুন খবর আনার।”